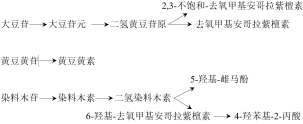
Soy isoflavones are a class of natural secondary metabolites with phytoestrogenic activity found in soybeans, primarily classified into free-type and conjugated-type, with the conjugated form accounting for 97%–98% of the total. This review summarizes the absorption, transformation, and metabolic pathways of soy isoflavones in the small intestine, liver, and large intestine of chickens. Conjugated isoflavones must be hydrolyzed by gut microbiota-secreted β-glucosidase into free aglycones before absorption. Free isoflavones undergo phase I and phase II metabolism in the liver, with some metabolites excreted into the intestine via bile, forming enterohepatic circulation. The unabsorbed fraction enters the large intestine and is further converted into highly active metabolites such as equol. Gut microbiota-mediated hydrolysis and conversion are key to producing these bioactive metabolites. Application studies in poultry production demonstrate that soy isoflavones can effectively improve laying and growth performance, enhance meat quality, and boost antioxidant capacity, immune regulation, and intestinal health by modulating endocrine function, activating antioxidant and immune-related signaling pathways (e.g., NF-κB and MAPK), and regulating gut microbiota. This review systematically summarizes the multifaceted biological effects and underlying mechanisms of soy isoflavones, providing a theoretical basis for their rational and standardized application in livestock and poultry farming.
| Published in | Science Research (Volume 14, Issue 3) |
| DOI | 10.11648/j.sr.20261403.16 |
| Page(s) | 105-112 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2026. Published by Science Publishing Group |
Soy Isoflavones, Chicken, Metabolism, Gut Microbiota, Free Type
大类 | 小类 | 中文名1 | 中文别名2 | 英文名1 | 英文别名2 | 分子式 | 结构式 |
|---|---|---|---|---|---|---|---|
游离型 | 大豆苷元 | 大豆甙元、黄豆苷元、大豆素、4, 7-二羟基异黄酮 | Daidzein | Daidsein、Daidzeol、4',7-Dihydroxy isoflavone | C15H10O4 |
| |
染料木素 | 染料木黄酮、金雀异黄素、4',5,7-三羟基异黄酮 | Genistein | Genisteol、 4',5,7-trihydroxy isoflavone | C15H10O5 |
| ||
黄豆黄素 | 4',7-二羟基-6-甲氧基异黄酮 | Glycitein | 4',7-Dihydroxy-6-methoxy isoflavone | C16H12O5 |
| ||
结合型 | 葡萄糖苷型 | 大豆苷 | 大豆甙、黄豆苷、异黄酮苷、7-羟基-3-(4-羟苯基)-异黄酮-7-糖苷 | Daidzin | Daidzoside、 daidzein 7-O-β-D-glucoside Daidzein-7-glucoside | C21H20O9 |
|
染料木苷 | 染料木甙、4',5,7-三羟异黄酮-7-糖苷 | Genistin | Genistein glucoside、 4',5,7-trihydroxyisoflavone-7-D-glucoside | C21H20O10 |
| ||
黄豆黄苷 | 黄豆黄甙、大豆黄苷、大豆黄甙 | Glycitin | Glycitein 7-O-glucoside; | C22H22O10 |
| ||
乙酰基 葡萄糖苷型 | 乙酰基黄豆苷 | 6"-O-乙酰黄豆苷 | Daidzein 6''-O-Acetate | 6''-o-acetyl daidzin | C23H22O10 |
| |
乙酰基染料木苷 | 6′′-O-乙酰染料木苷 | Genistin 6''-O-Acetate | 6''-o-acetyl genistin | C23H22O11 |
| ||
乙酰基黄豆黄苷 | 6''-O-乙酰黄豆黄苷 | Glycitin 6''-O-Acetate | 6''-o-acetyl glycitin | C24H24O11 |
| ||
丙二酰基 葡萄糖苷型 | 丙二酰基大豆苷 | 6''-O-丙二酰基大豆苷 | Daidzin 6''-O-Malonate | 6"-O-Malonyl Daidzin、malonyl daidzin | C24H22O12 |
| |
丙二酰基染料木苷 | 丙二酰基染料木甙 | Genistin 6''-O-Malonate | 6''-o-malonylgenistin 、Genistin malonate | C24H22O13 |
| ||
丙二酰基黄豆黄苷 | 丙二酰黄豆黄甙、6''-O-丙二酰黄豆黄苷 | Glycitin 6''-O-Malonate | 6"-O-Malonyl Glycitin | C25H24O13 |
|
| [1] | 何灵芝, 覃圣, 撒瑞雪, 徐硕, 程香, 马俊贤. 大豆异黄酮对生殖细胞增殖影响的研究进展 [J]. 现代畜牧兽医, 2019, (12): 54-58. |
| [2] | Villares, A., Rostagno, M. A., Ana García-Lafuente, Eva Guillamón, & J. Alfredo Martínez. Content and profile of isoflavones in soy-based foods as a function of the production process. Food & Bioprocess Technology, 2011, 4(1), 27-38. |
| [3] | 李参参, 张淦, 常徽. 微生物发酵法转化大豆异黄酮研究进展 [J]. 食品研究与开发, 2023, 44(12): 218-224. |
| [4] | Xu J, Chen H, Li S. Understanding the molecular mechanisms of the interplay between herbal medicines and gut microbiota [J]. Medicinal Research Reviews, 2017, 37(5): 1140-1185. |
| [5] | 陈嘉序, 陈如扬, 连媛, 姚欣硕, 黄钰昕, 高瑶, 肖愈. 大豆异黄酮的生物转化及功能活性研究进展 [J]. 食品研究与开发, 2023, 42(9): 176-182. |
| [6] | 梁文欧, 赵力超, 方祥, 王丽. 大豆异黄酮与肠道微生物相互作用研究进展 [J]. 食品科学, 2019, 40(9): 283-288. |
| [7] | Fang W, Song R, Zhang X, Zhang X, Zhang X, Wang X, Fang Z, Xiao Y. Characterization of a Novel β-Glucosidase from Gongronella sp. W5 and Its Application in the Hydrolysis of Soybean Isoflavone Glycosides [J]. Journal of agricultural and food chemistry, 2014, 62(48): 11688-11695. |
| [8] | 鲍捷. 产β-葡萄糖苷酶乳酸菌的筛选及发酵豆浆中大豆异黄酮糖苷代谢研究 [D]. 大连: 大连工业大学, 2023. |
| [9] | 王丹媚.β-葡萄糖苷酶水解大豆异黄酮及其在提高土鸡品质中的探究 [D]. 金华: 浙江师范大学, 2022. |
| [10] | 周文红, 郭咪咪, 李秀娟, 毕艳红, 王朝宇, 段章群. 大豆异黄酮提取及其生物转化的研究进展 [J]. 粮油食品科技, 2019, 27(5): 37-42. |
| [11] | 赵慧颖, 余诗强, 蒋林树, 熊本海. 大豆异黄酮的代谢及其对动物肠道保护机制的研究进展 [J]. 动物营养学报, 2022, 34(7): 4132-4142. |
| [12] | Snchez-calvo J M, Rodr guez-iglesias M A, Molinillo J M G, Macias F A. Soy isoflavones and their relationship with microflora: beneficial effects on human health in equol producers [J]. Phytochemistry Reviews, 2013, 12(4): 979-1000. |
| [13] | Islam M A, Punt A, Spenkelink B, et al. Murk A J, Rolaf van Leeuwen F X, Rietjens I M C M. Conversion of major soy isoflavone glucosides and aglycones in in vitro intestinal models [J]. Molecular nutrition & food research, 2014, 58(3): 503-515. |
| [14] | Rowland I, Faughnan M, Hoey L, Wähälä K, Williamson G, Cassidy A. Bioavailability of phyto⁃oestrogens [J]. British Journal of Nutrition, 2003, 89(Suppl.1): S45-S58. |
| [15] | Wang X, Ma Y, Wu X, Chen B, Fang X, Zhong Q, Liao Z, Wang J, Wang L. The dietary intervention of synbiotic preparation promotes the bioconversion of soy isoflavones to equol and its metabolic mechanism [J]. Journal of Functional Foods, 2023, 109(000): 15. |
| [16] | Barnes S, Prasain J, D’alessandro T, Arabshahi A, Botting N, Lila M A, Jackson G, Janle E M, Weaver C M. The metabolism and analysis of isoflavones and otherdietary polyphenols in foods and biological systems [J]. Food & Function, 2011, 2(5): 235-244. |
| [17] | Maruo T, Sakamoto M, Ito C, Toda T; Benno Y. Adlercreutzia equolifaciens gen. nov., sp. nov., an equol-producing bacterium isolated from human faeces, and emended description of the genus Eggerthella [J]. International Journal of Systematic & Evolutionary Microbiology, 2008, 58(5): 1221-1227. |
| [18] | Rüfer C E, Kulling S E. Antioxidant activity of isoflavones and their major metabolites using different in vitro assays [J]. Journal of Agricultural & Food Chemistry, 2006, 54(8): 2926-2931. |
| [19] | 李燕, 蒋守群, 陈伟. 雌马酚的产生、生物学功能及潜在应用 [J]. 动物营养学报, 2014, 26(11): 3236-3244. |
| [20] | Xie Y, Liu Z, Gao Y, Wang X, Hao Q, Yu X. Bioconversion of genistein to(–)-5-hydroxy-equol by a newly isolated cock intestinal anaerobic bacterium [J]. Journal of Chinese Pharmaceutical Sciences, 2015, 24 (7), 442–448. |
| [21] | 王秀伶, 王烨. 膳食中的植物雌激素、肠道菌群与人类健康 [J]. 世界华人消化杂志, 2016, (35): 51-67. |
| [22] | Lee D, Kim M J, Ahn J, Lee S H, Lee H, Kim J H, Park S H, Jang Y J, Ha TY, Jung CH. Nutrikinetics of isoflavone metabolites after fermented soybean product (cheonggukjang) ingestion in ovariectomized mice [J]. Molecular Nutrition & Food Research, 2017, 61(12): 1700322. |
| [23] | Obianwuna U E, Chang X, Wu R, Wang J, Zhang H J, Wu S G, Qiu K. 2025. Effect of Genistein and Glycitein on production performance, egg quality, antioxidant function, reproductive hormones and related-genes in pre-peak laying hens [J]. Poultry science, 103(10): 103952. |
| [24] | 强桃艳. 2024. 大豆异黄酮对不同产蛋水平产蛋后期蛋鸡生产性能、蛋品质和卵巢功能的影响 [D]. 成都: 四川农业大学. |
| [25] | 王贝贝, 武书庚, 张海华, 张海军, 郝二英, 邱凯. 饲粮添加大豆异黄酮对产蛋后期蛋鸡生产的影响 [J]. 畜牧兽医学报, 2025, 56(1): 295-306. |
| [26] | 林厦菁, 苟钟勇, 范秋丽, 叶金玲, 王一冰, 蒋守群. 大豆异黄酮对黄羽肉种母鸡产蛋、繁殖性能及血浆生化指标的影响 [J].中国畜牧兽医, 2023, 50(11): 4454-4462. |
| [27] | Zhao R Q, Zhou Y C, Ni Y D, Lu L Z, Tao Z R, Chen W H. Effect of daidzein on egg-laying performance in Shaoring duck breeders during different stages of the egg production cycle [J]. British Poultry Science, 2005, 46: 175-181. |
| [28] | 曹满湖, 罗理成, 孙佳静, 方热军, 朱静波. 大豆异黄酮对产蛋后期蛋鸡卵巢机能的影响 [J]. 动物营养学报, 2016, 28(8): 7. |
| [29] | 吴晶, 童海洋, 魏晓静, 倪迎冬. 产蛋后期母鸡日粮中添加大豆异黄酮对产蛋率及肝脏相关基因表达的影响 [C]. 全国动物生理生化第十一次学术交流会, 2025. |
| [30] | 田何芳, 金永燕, 庄智威, 戴振宇, 刘文超, 郑春田, 张亚男. 大豆异黄酮对产蛋后期京粉1号蛋鸡产蛋性能、蛋品质、血浆生化指标和抗氧化能力的影响 [J]. 中国家禽, 2021, 43(12): 6. |
| [31] | Lv Z, Fan H, Zhang B, Xing K, Guo Y. Dietary genistein supplementation for breeders and their offspring improves the growth performance and immune function of broilers [J]. Scientific Reports, 2018, 8(1): 5161. |
| [32] | Fan H, Lv Z P, Gan L P, Ning C, Zhui Li,Yang M H, Zhang B B, Song B C, Li G, Tang D Z, Gao J X, Yan S J, Wang Y L, Liu J F, Guo Y M.. A Novel lncRNA regulates the toll-like receptor signaling pathway and related immune function by stabilizing fos mRNA as a competitive endogenous RNA [J]. Frontiers in immunology, 10: 838-838. |
| [33] | 刘帅, 熊云霞, 孙铝辉, 王丽.大豆异黄酮在猪禽养殖中应用的研究进展 [J]. 动物营养学报, 2021, 33(4): 1822-1831. |
| [34] | 许啸, 齐智利. 大豆异黄酮对畜禽生理机能的调控 [J]. 动物营养学报, 2012, 24(3): 436-438. |
| [35] | Bodine S C, Stitt T N, Gonzalez M, Kline W O, Yancopoulos G D. Akt/mTOR pathway is a crucial regulator of skeletal muscle hypertrophy and can prevent muscle atrophy in vivo [J]. Nat Cell Biol, 2001, 3(11): 1014-1019. |
| [36] | Cederroth C R, Vinciguerra M, Gjinovci A, Franoise Kü, Nef S. Dietary phytoestrogens activate AMP-activated protein kinase with improvement in lipid and glucose metabolism [J]. Diabetes, 2008, 57(5): 1176-1185. |
| [37] | Mueller M, Lukas B, Novak J, Simoncini T, Genazzani A R, Jungbauer A. Oregano: a source for peroxisome proliferator-activated receptor gamma antagonists [J]. J Agric Food Chem, 2008, 56(24): 11621-11630. |
| [38] | 王冬生, 韩婧, 康文博, 赵建宁. 植物雌激素防治骨质疏松作用的机制进展 [J]. 中国骨质疏松杂志, 2016, 22(5): 632-640. |
| [39] | Utami M M D, Hertamawati R T. Dietary edamame soybean isoflavon concentrate on improving carcass quality of broilers [J]. IOP Conference Series: Earth and Environmental Science, 2020, 411(1): 012035. |
| [40] | Kamboh A A, Zhu W Y. Individual and combined effects of genistein and hesperidin supplementation on meat quality in meat type broiler chickens [J]. J Sci Food Agric, 2013, 93(13): 3362-3367. |
| [41] | Payne R L, Bidner T D, Southern L L, Geaghan J P. Effects of dietary soy isoflavones on growth, carcass traits, and meat quality in growing-finishing pigs [J]. J Anim Sci, 2001, 79(5): 1230-1239. |
| [42] | 官丽辉, 马旭平, 刘海斌, 张立永, 关琛, 王志伟. 大豆异黄酮对坝上长尾鸡卵巢功能、生殖激素和肌肉品质的影响 [J]. 中国兽医学报, 2021, 41(2): 338-344. |
| [43] | 叶金玲, 范秋丽, 王一冰, 林厦菁, 阮栋, 罗琦丽, 苟钟勇, 蒋守群. 磷脂与大豆异黄酮对黄羽肉鸡生长性能、肉品质及抗氧化功能的影响 [J]. 中国畜牧兽医, 2024, 51(5): 1857-1867. |
| [44] | 刘皙洁, 张桂春, 张维生. 半胱胺、大豆黄酮对肉仔鸡脂肪代谢的影响 [J]. 东北农业大学学报, 2003, 34(2): 171-175. |
| [45] | 宋凯. 大豆异黄酮的生理功能及其对畜产品品质的影响 [J]. 饲料博览, 2008, (7): 5-7. |
| [46] | 陈浩瀚, 王锦湘, 王敏奇. 大豆异黄酮对生长猪生长性能、血清生化、免疫和抗氧化能力的影响 [J]. 中国畜牧杂志, 2022, 58(5): 229-238. |
| [47] | Jiang Z Y, Zhou G L, Lin Y C, Jiang S Q, Zheng C T, Han L M, Chen F. Effects of soybean isoflavones on in vitro antioxidative capacity of satellite cells of porcine skeletal muscles [J]. Agr Sci China, 2011, 10(1): 120-125. |
| [48] | Shimada Y, Yasuda S, Takahashi M, Hayashi T, Miyazawa N, Sato I, Abiru Y, Uchiyama S, Hishigaki H. Cloning and expression of a novel nadp(h)-dependent daidzein reductase, an enzyme involved in the metabolism of daidzein, from equol-producing lactococcus strain 20-92 [J]. Applied and Environmental Microbiology, 2010, 76(17), 5892-5901. |
| [49] | Gun-Ae, Y, Sunmin P. Antioxidant action of soy isoflavones on oxidative stress and antioxidant enzyme activities in exercised rats [J]. Nutrition Research and Practice, 2014, 8(6): 618-624. |
| [50] | 杨帆.大豆异黄酮的生理作用及其在畜牧生产中的应用 [J]. 饲料与畜牧: 新饲料, 2010, (2): 7. |
| [51] | Chadha R, Bhalla Y, Jain A, Chadha K, Karan M. Dietary soy isoflavone: a mechanistic insight [J]. Natural Product Communications, 2017, 12(4); 1069-1074. |
| [52] | 吕增鹏. 2018. 染料木黄酮调节种鸡及其子代脂代谢与免疫的分子机理研究 [D]. 北京: 中国农业大学. |
| [53] | 王洋, 孙玉刚. 大豆异黄酮片对肉仔公鸡免疫功能的影响 [J]. 中国农学通报, 2009, 25(20): 35-37. |
| [54] | Sakai T, Kogiso M. Soy isoflavones and immunity [J]. J Med Invest, 2008, 55(3-4): 167-173. |
| [55] | 付丽佳, 林海, 王玉华.大豆异黄酮雌激素样作用的研究进展 [J]. 中国伤残医学, 2008, 16(6): 2. |
| [56] | Salces F R, Rostagno M A, Farfan J A. Novel process of hydration, followed by incubation and thermal processing, for high isoflavone bioconversion in soybeans [J]. Food Research International, 2019, 121(JUL.): 691-696. |
| [57] | 任维利. 大豆异黄酮的作用研究进展 [J]. 食品安全导刊, 2021, 14: 74-75. |
| [58] | Hou Q, Huang J, Zhao L, et al. Dietary genistein increases microbiota-derived short chain fatty acid levels, modulates homeostasis of the aging gut, and extends healthspan and lifespan [J]. Pharmacological Research, 2023. |
| [59] | Jeong J W, Lee H H, Han M H, Kim G Y, Kim W J, Choi Y H. Anti-inflammatory effects of genistein via suppression of the tolllike receptor 4-mediated signaling pathway in lipopolysaccharide-stimulated BV2 microglia [J]. Chemico Biological Interactions, 2014, 212: 30-39. |
| [60] | Mccarty M F, Lerner A. Perspective: Prospects for nutraceutical support of intestinal barrier function [J]. Advances in Nutrition, 2020, 12(3). |
| [61] | 程东静. 2019. 大豆异黄酮对高脂肥胖大鼠结肠屏障功能的影响 [D]. 雅安: 四川农业大学. |
| [62] | Vázquez L, Flórez A B, Guadamuro L, Mayo B. Effect of soy isoflavones on growth of representative bacterial species from the human gut [J]. Nutrients, 2017, 9(7), 727. |
| [63] | Hong H, Landauer M R, Foriska M A, Ledney G D. Antibacterial activity of the soy isoflavone genistein [J]. Journal of Basic Microbiology, 2006, 46(4): 329-335. |
| [64] | 王海涛. 2009. 大豆异黄酮的抑菌活性及其机制的研究 [D].硕士学位论文.大连: 辽宁师范大学. |
| [65] | 刘佳明, 吕德志, 张大伟. 双歧杆菌发酵对豆芽汁中大豆异黄酮含量及组分影响的研究 [J]. 中国微生态学杂志, 2004, 16(4): 213-214. |
| [66] | Mika A, Greenwood B N, Chichlowski M, Borchert D, Hulen K A, Berg B M, Paton M, Fleshner M. Dietary prebiotics increase Bifidobacterium spp. and Lactobacillus spp. in the gut and promote stress resistance [J]. Brain Behavior & Immunity, 2014, 40: e45. |
APA Style
Sun, B., Ma, Y., Li, Y., Bie, J., Song, D. (2026). Soy Isoflavones in Poultry Production: Research Advances on Metabolism, Functions, and Applications. Science Research, 14(3), 105-112. https://doi.org/10.11648/j.sr.20261403.16
ACS Style
Sun, B.; Ma, Y.; Li, Y.; Bie, J.; Song, D. Soy Isoflavones in Poultry Production: Research Advances on Metabolism, Functions, and Applications. Sci. Res. 2026, 14(3), 105-112. doi: 10.11648/j.sr.20261403.16
@article{10.11648/j.sr.20261403.16,
author = {Baosheng Sun and Yujiao Ma and Yan Li and Junyan Bie and Danni Song},
title = {Soy Isoflavones in Poultry Production: Research Advances on Metabolism, Functions, and Applications},
journal = {Science Research},
volume = {14},
number = {3},
pages = {105-112},
doi = {10.11648/j.sr.20261403.16},
url = {https://doi.org/10.11648/j.sr.20261403.16},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.sr.20261403.16},
abstract = {Soy isoflavones are a class of natural secondary metabolites with phytoestrogenic activity found in soybeans, primarily classified into free-type and conjugated-type, with the conjugated form accounting for 97%–98% of the total. This review summarizes the absorption, transformation, and metabolic pathways of soy isoflavones in the small intestine, liver, and large intestine of chickens. Conjugated isoflavones must be hydrolyzed by gut microbiota-secreted β-glucosidase into free aglycones before absorption. Free isoflavones undergo phase I and phase II metabolism in the liver, with some metabolites excreted into the intestine via bile, forming enterohepatic circulation. The unabsorbed fraction enters the large intestine and is further converted into highly active metabolites such as equol. Gut microbiota-mediated hydrolysis and conversion are key to producing these bioactive metabolites. Application studies in poultry production demonstrate that soy isoflavones can effectively improve laying and growth performance, enhance meat quality, and boost antioxidant capacity, immune regulation, and intestinal health by modulating endocrine function, activating antioxidant and immune-related signaling pathways (e.g., NF-κB and MAPK), and regulating gut microbiota. This review systematically summarizes the multifaceted biological effects and underlying mechanisms of soy isoflavones, providing a theoretical basis for their rational and standardized application in livestock and poultry farming.},
year = {2026}
}
TY - JOUR T1 - Soy Isoflavones in Poultry Production: Research Advances on Metabolism, Functions, and Applications AU - Baosheng Sun AU - Yujiao Ma AU - Yan Li AU - Junyan Bie AU - Danni Song Y1 - 2026/06/09 PY - 2026 N1 - https://doi.org/10.11648/j.sr.20261403.16 DO - 10.11648/j.sr.20261403.16 T2 - Science Research JF - Science Research JO - Science Research SP - 105 EP - 112 PB - Science Publishing Group SN - 2329-0927 UR - https://doi.org/10.11648/j.sr.20261403.16 AB - Soy isoflavones are a class of natural secondary metabolites with phytoestrogenic activity found in soybeans, primarily classified into free-type and conjugated-type, with the conjugated form accounting for 97%–98% of the total. This review summarizes the absorption, transformation, and metabolic pathways of soy isoflavones in the small intestine, liver, and large intestine of chickens. Conjugated isoflavones must be hydrolyzed by gut microbiota-secreted β-glucosidase into free aglycones before absorption. Free isoflavones undergo phase I and phase II metabolism in the liver, with some metabolites excreted into the intestine via bile, forming enterohepatic circulation. The unabsorbed fraction enters the large intestine and is further converted into highly active metabolites such as equol. Gut microbiota-mediated hydrolysis and conversion are key to producing these bioactive metabolites. Application studies in poultry production demonstrate that soy isoflavones can effectively improve laying and growth performance, enhance meat quality, and boost antioxidant capacity, immune regulation, and intestinal health by modulating endocrine function, activating antioxidant and immune-related signaling pathways (e.g., NF-κB and MAPK), and regulating gut microbiota. This review systematically summarizes the multifaceted biological effects and underlying mechanisms of soy isoflavones, providing a theoretical basis for their rational and standardized application in livestock and poultry farming. VL - 14 IS - 3 ER -
Hubei Jingruitianheng Biotechnology Co., Ltd., Yichang, China; College of Animal Science and Technology, China Agricultural University, Beijing, China
Hubei Jingruitianheng Biotechnology Co., Ltd., Yichang, China
Hubei Jingruitianheng Biotechnology Co., Ltd., Yichang, China
Hubei Jingruitianheng Biotechnology Co., Ltd., Yichang, China
Hubei Jingruitianheng Biotechnology Co., Ltd., Yichang, China
Information