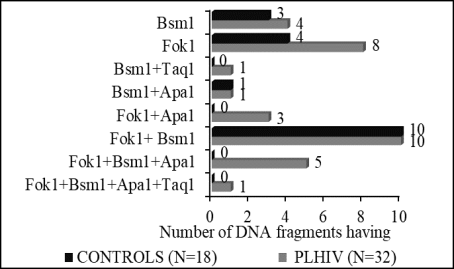
Introduction: Studies conducted in Côte d'Ivoire with HIV patients have revealed disorders in phosphorus/calcium metabolism and 25-hydroxyvitmin D3 status. Due to the important role played by VDR-1,25 (OH)2 D3 in the biological activity of 25-hydroxyvitamin D3, it has become necessary to describe the polymorphism of 4 fragments (Fok-1, Bsm-1, Apa-1, Taq-1) of the nuclear receptor gene Vitamin D3 (VDR) in PLHIV. Methods: The study collected blood samples from 50 individuals, including 32 HIV positive patients and 18 HIV negative as a control. The extraction of genomic DNA from peripheral blood mononuclear cells (PBMC) was done using the QIAamp kit (QIAGEN). After amplification, the PCR products obtained were purified using the "High Pure PCR product purification kit" and sequenced at Eurofilms MWG operon. Results: The prevalence of the Fok-1 mutant C allele was 87.5% (28/32) in PLHIV against 77.8% (14/18) in controls. The single mutant genotype Thr1Met4 was 62.5% in PLHIV versus 55.6% in controls. The prevalence of the mutant allele A (g.63510 A) of Bsm-1 (rs 1544410) was 28% in HIV-infected against 66.7% (12/18) in controls. The mutant GG genotype was more prevalent in 34.4% of PLHIV. Two types of mutant alleles A (g.65023 A) and C (g.65024 C) and the AC mutant genotype of Apa-1 (rs 7975232) were more observed in 28% of those infected with HIV versus 0% in controls. No significant difference was observed between the frequencies of wild T (g.65058 T) allele and mutant C or A (g.65058 C or A) of Taq-1 (rs 731236) (p = 0.47). The wild genotype Ile352 was the most common regarding the Taq-1 polymorphism. Ten PLHIV and controls have the combination Fok-1 + Bsm-1 in common. The combinations Fok-1 + Bsm-1 + Apa-1 and Fok-1 + Apa-1 were specific to PLHIV. Conclusion: The blood sample of PLHIV subjects is characterized by polymorphisms affecting Fok-1, Bsm-1, and Apa-1. In addition, the combination of Fok-1 and Bsm-1 was observed in both PLHIV patients and controls.
| Published in | Biochemistry and Molecular Biology (Volume 10, Issue 4) |
| DOI | 10.11648/j.bmb.20251004.11 |
| Page(s) | 61-70 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2025. Published by Science Publishing Group |
Polymorphism, VDR Gene, HIV Positive Patients, Côte d'Ivoire
Fragment analysed | Position analysed | Sample PLHIV n (%) | Sample Control n (%) | P value |
|---|---|---|---|---|
Fok-1 Exon2 | Alleles | |||
g.30920 T | 4 (12.5) | 4 (22.2) | 0.62 | |
g.30920 C | 28 (87.5) | 14 (77.8) | 0.62 | |
g.30929 T | 24 (75) | 14 (77.8) | 0.86 | |
g.30929 C | 8 (25) | 4 (22.2) | 1 | |
Genotypes | ||||
Met1Met4 (Wild) | 4 (12.5) | 4 (22.2) | 0.62 | |
Thr1Met4 (Simple mutant) | 20 (62.5) | 10 (55.6) | 0.86 | |
Thr1Thr4 (Double mutant) | 8 (25) | 4 (22.2) | 1 | |
Fragment analysed | Position analysed | Sample PLHIV n (%) | Sample Control n (%) | p value |
|---|---|---|---|---|
Bsm-1 in intron 8 | Alleles | |||
g.63510 G | 23 (71.9) | 6 (33.3) | 0.02 | |
g.63510 A | 9 (28.1) | 12 (66.7) | 0.02 | |
g.63905 A | 12 (37.5) | 6 (33.3) | 1 | |
g.63905 G | 17 (53.1) | 8 (44.4) | 0.77 | |
g.63905 C | 3 (9.4) | 4 (22.2) | 0.4 | |
Genotypes | ||||
GA | 11 (34.4) | 4 (22.2 | 0.56 | |
GG | 11 (34.4) | 1 (5.6) | 0.05 | |
AG | 6 (18.7) | 7 (38.9) | 0.22 | |
AC | 2 (6.2) | 3 (16.7) | 1 | |
AA | 1 (3.1) | 2 (11.1) | 0.6 | |
GC | 1 (3.1) | 1 (5.6) | 1 | |
Fragment analysed | Position analysed | Sample PLHIV n (%) | Sample Control n (%) | p value |
|---|---|---|---|---|
Apa-1 in intron 8 | Alleles | |||
g.65023 C | 23 (71.9) | 18 (100) | 0.03 | |
g.65023 A | 9 (28.1) | 0 (0) | 0.03 | |
g.65024 A | 23 (71.9) | 18 (100) | 0.03 | |
g.65024 C | 9 (28.1) | 0 (0) | 0.03 | |
Genotypes | ||||
CA | 23 (71.9) | 18 (100) | 0.03 | |
AC | 9 (28.1) | 0 (0) | 0.03 | |
Fragment analysed | Position analysed | Sample PLHIV n (%) | Sample Control n (%) | P value |
|---|---|---|---|---|
Taq-1 in exon 9 | Alleles | |||
g.65058 T | 19 (59.4) | 6 (33.3) | 0.14 | |
g.65058 A | 13 (40.6) | 10 (55.6) | 0.47 | |
g.65058 C | 0 (0) | 2 (11.1) | 0.24 | |
Genotypes | ||||
Ile352 (Wild) | 30 (93.7) | 17 (94.4) | 1 | |
Leu352 (Simple mutant) | 2 (6.2) | 1 (5.6) | 1 | |
VDR | Vitamin D nuclear Receptor |
PLHIV | People Living with HIV |
PBMC | Peripheral Blood Mononuclear Cells |
SNP | Single Nucleotide Polymorphism |
| [1] | Alqudah, M., Yaseen, M., Yaseen, M. HIV-1 strategies to overcome the immune system by evading and invading innate immune system. HIV & AIDS Review. International Journal of HIV-Related Problems, 2016, 15(1), 1-12. |
| [2] | TPE ADVIH: HIV self-testing in Côte d’Ivoire: nationwide roll-out of an innovative tool to improve access to testing for most-at-risk populations. (accessed April 3, 2025) |
| [3] | Shin, D. H., Martinez, S. S., Parsons, M., Jayaweera, D. T., Campa, A., Baum, M. K. Relationship of Oxidative Stress with HIV Disease Progression in HIV/HCV Co-infected and HIV Mono-infected Adults in Miami. International journal of bioscience, biochemistry and bioinformatics, 2012, 2(3), 217–223. |
| [4] | Konings E. J., Committee on Food Nutrition. Water-soluble vitamins. Journal of AOAC International, 2006, 89(1), 285–288. |
| [5] | Andrés, C. M. C., Pérez de la Lastra, J. M., Juan, C. A., Plou F. J., Pérez-Lebeña E. Antioxidant Metabolism Pathways in Vitamins, Polyphenols, and Selenium: Parallels and Divergences. International Journal of Molecular Sciences, 2024, 25(5), 2600. |
| [6] | Gil, Á., Plaza-Diaz, J., Mesa, M. D. Vitamin D: Classic and Novel Actions. Annals of nutrition & metabolism, 2018, 72(2), 87–95. |
| [7] | Larriba, M. J., González-Sancho, J. M., Barbáchano, A., Niell, N., Ferrer-Mayorga, G., Muñoz, A. Vitamin D Is a Multilevel Repressor of Wnt/b-Catenin Signaling in Cancer Cells. Cancers, 2013, 5(4), 1242–1260. |
| [8] | Nieto, G., Barber, Y., Rubio, M. C., Rubio, M., Fibla, J. Association between AIDS disease progression rates and the Fok-I polymorphism of the VDR gene in a cohort of HIV-1 seropositive patients. The Journal of steroid biochemistry and molecular biology, 2004, 89-90(1-5), 199–207. |
| [9] | Gichuhi, C., Kariuki, D., Nyerere, A., Riyat, M. Studies on Vitamin D Levels in Serum of HIV Infected Patients: Their Effect on Progression towards AIDS. World Journal of AIDS, 2014, 4(4), 422-429. |
| [10] | de la Torre, M. S., Torres, C., Nieto, G., Vergara, S., Carrero, A. J., Macías, J., Pineda, J. A., Caruz, A., Fibla, J. Vitamin D receptor gene haplotypes and susceptibility to HIV-1 infection in injection drug users. The Journal of infectious diseases, 2008, 197(3), 405–410. |
| [11] | Małodobra-Mazur, M., Paduch, A., Lebioda, A., Konopacka, M., Rogoliński, J., Szymczyk, C., Wierzgoń, J., Maciejewski, A., Chmielik, E., Jonkisz, A., Półtorak, S., Dobosz, T. VDR gene single nucleotide polymorphisms and their association with risk of oral cavity carcinoma. Acta biochimica Polonica, 2012, 59(4), 627–630. |
| [12] | Belorusova, A. Y., Rochel, N. Structural Studies of Vitamin D Nuclear Receptor Ligand-Binding Properties. Vitamins and hormones, 2016, 100, 83–116. |
| [13] | Uitterlinden, A. G., Fang, Y., Van Meurs, J. B., Pols, H. A., Van Leeuwen, J. P.. Genetics and biology of vitamin D receptor polymorphisms. Gene, 2004, 338(2), 143–156. |
| [14] | Köstner, K., Denzer, N., Müller, C. S., Klein, R., Tilgen, W., Reichrath, J. The relevance of vitamin D receptor (VDR) gene polymorphisms for cancer: a review of the literature. Anticancer research, 2009, 29(9), 3511–3536. |
| [15] | Tombari, S., Amri, Y., Hasni, Y., Hadj Fredj, S., Salem, Y., Ferchichi, S., Essaddam, L., Messaoud, T., Dabboubi, R. Vitamin D status and VDR gene polymorphisms in patients with growth hormone deficiency: A case control Tunisian study. Heliyon, 2024, 10(14), e34947. |
| [16] | Panierakis, C., Goulielmos, G., Mamoulakis, D., Petraki, E., Papavasiliou, E., Galanakis, E. Vitamin D receptor gene polymorphisms and susceptibility to type 1 diabetes in Crete, Greece. Clinical immunology (Orlando, Fla.), 2009, 133(2), 276–281. |
| [17] | Ghosn, J., Viard, J. P. Vitamine D et infections [Vitamin D and infectious diseases]. Presse medicale (Paris, France: 1983), 2013, 42(10), 1371–1376. |
| [18] | Beard, J. A., Bearden, A., Striker, R. Vitamin D and the anti-viral state. Journal of clinical virology: the official publication of the Pan American Society for Clinical Virology, 2011, 50(3), 194–200. |
| [19] | Boyvin, L, Aké, J. A., Séri, K. L., M’boh, G. M., Yapo, A. F., Djaman J. A. 25 (OH) Vitamin D level and Calcium/Phosphorus Metabolism Disorders in Patients Living with HIV in Abidjan. International Journal of Biochemistry Research & Review, 2017, 17(4), 1–7. |
| [20] | Aké, A. J. A., Boyvin, L., Séri, K. L., Allico, M. J. M., Yéo, K. D., Yapo, A. F., Djaman, A. J. Vitamin D Deficiency and Evaluation of the Parathyroid Hormone Status in People Living with HIV in Côte d’Ivoire. Asian Journal of Research in Biochemistry, 2018, 2(1), 1–10. |
| [21] | Pawlowska, E., Wysokinski, D., Blasiak, J. Nucleotide Excision Repair and Vitamin D--Relevance for Skin Cancer Therapy. International journal of molecular sciences, 2016, 17(4), 372. |
| [22] | Crucitti, T., Taylor, D., Beelaert, G., Fransen, K., Van Damme, L. Performance of a rapid and simple HIV testing algorithm in a multicenter phase III microbicide clinical trial. Clinical and vaccine immunology: CVI, 2011, 18(9), 1480–1485. |
| [23] | Sagar N. A., Sen M., Yadav, V. K. Serodiagnosis of HIV by Rapid Test and ELISA Test Assay in a Tertiary Care Centre in Northern India. International Journal of Current Microbiology and Applied Sciences, 2015, 4(10): 623-629 |
| [24] | Methods in Molecular Medicine. In Molecular Pathology Protocols. Edited by: Anthony A. Killeen. Humana Totowa, NJ. 2001, XIV, 491 p. |
| [25] | Chakraborty, A., Mishra, A. K., Soni, A., Regina, T., Mohil, R., Bhatnagar, D., Bhatnagar, A., Chintamani, C., Sharma, P. C., Saxena, S. Vitamin D receptor gene polymorphism(s) and breast cancer risk in north Indians. Cancer detection and prevention, 2009, 32(5-6), 386–394. |
| [26] | Rashedi, J., Asgharzadeh, M., Moaddab, S. R., Sahebi, L., Khalili, M., Mazani, M., Abdolalizadeh, J. Vitamin d receptor gene polymorphism and vitamin d plasma concentration: correlation with susceptibility to tuberculosis. Advanced pharmaceutical bulletin, 2014, 4(Suppl 2), 607–611. |
| [27] | Ashktorab, H., Nguza, B., Fatemi, M., Nouraie, M., Smoot, D. T., Schäffer, A. A., Kupfer, S. S., Camargo, C. A., Jr, Brim, H. Case-control study of vitamin D, dickkopf homolog 1 (DKK1) gene methylation, VDR gene polymorphism and the risk of colon adenoma in African Americans. PloS one, 2011, 6(10), e25314. |
| [28] | O Neill, V., Asani, F. F., Jeffery, T. J., Saccone, D. S., Bornman, L. Vitamin D Receptor Gene Expression and Function in a South African Population: Ethnicity, Vitamin D and FokI. PloS one, 2013, 8(6), e67663. |
| [29] | Neela, V. S., Suryadevara, N. C., Shinde, V. G., Pydi, S. S., Jain, S., Jonnalagada, S., Singh, S. S., Valluri, V. L., Anandaraj, M. P. Association of Taq I, Fok I and Apa I polymorphisms in Vitamin D Receptor (VDR) gene with leprosy. Human immunology, 2015, 76(6), 402–405. |
| [30] | Miyamoto, K., Kesterson, R. A., Yamamoto, H., Taketani, Y., Nishiwaki, E., Tatsumi, S., Inoue, Y., Morita, K., Takeda, E., Pike, J. W. Structural organization of the human vitamin D receptor chromosomal gene and its promoter. Molecular endocrinology (Baltimore, Md.), 1997, 11(8), 1165–1179. |
| [31] | Mahmoud A. A., Ali A. H. K. Vitamin D receptor gene polymorphism and 25 hydroxy vitamin D levels in Egyptian patients with pulmonary tuberculosis. Egypt J Chest Dis Tuberc, 2014, 63(3), 651-5. |
| [32] | Suneetha, P. V., Sarin, S. K., Goyal, A., Kumar, G. T., Shukla, D. K., Hissar, S. Association between vitamin D receptor, CCR5, TNF-alpha and TNF-beta gene polymorphisms and HBV infection and severity of liver disease. Journal of hepatology, 2006, 44(5), 856–863. |
| [33] | Larcombe, L. A., Orr, P. H., Lodge, A. M., Brown, J. S., Dembinski, I. J., Milligan, L. C., Larcombe, E. A., Martin, B. D., Nickerson, P. W. Functional gene polymorphisms in canadian aboriginal populations with high rates of tuberculosis. The Journal of infectious diseases, 2008, 198(8), 1175–1179. |
| [34] |
Zhang, L., Yin, X., Wang, J., Xu, D., Wang, Y., Yang, J., Tao, Y., Zhang, S., Feng, X., Yan, C. Associations between VDR Gene Polymorphisms and Osteoporosis Risk and Bone Mineral Density in Postmenopausal Women: A systematic review and Meta-Analysis. Scientific reports, 2018, 8(1), 981.
https://doi.org/10.1038/s41598-017-18670-7 (Retraction published Sci Rep, 2021, 11(1), 9030). |
| [35] | Alagarasu, K., Selvaraj, P., Swaminathan, S., Narendran, G., Narayanan, P. R. 5' regulatory and 3' untranslated region polymorphisms of vitamin D receptor gene in south Indian HIV and HIV-TB patients. Journal of clinical immunology, 2009, 29(2), 196–204. |
| [36] | McNamara, L., Takuva, S., Chirwa, T., MacPhail, P. Prevalence of common vitamin D receptor gene polymorphisms in HIV-infected and uninfected South Africans. International journal of molecular epidemiology and genetics, 2016, 7(1), 74–80. |
| [37] | Zmuda, J. M., Cauley, J. A., Ferrell, R. E. Molecular epidemiology of vitamin D receptor gene variants. Epidemiologic reviews, 2000, 22(2), 203–217. |
| [38] | Khan, M. I., Bielecka, Z. F., Najm, M. Z., Bartnik, E., Czarnecki, J. S., Czarnecka, A. M., Szczylik, C. Vitamin D receptor gene polymorphisms in breast and renal cancer: current state and future approaches (review). International journal of oncology, 2014, 44(2), 349–363. |
| [39] | Cao, Y., Wang, X., Cao, Z., Cheng, X. Association of Vitamin D receptor gene TaqI polymorphisms with tuberculosis susceptibility: a meta-analysis. International journal of clinical and experimental medicine, 2015, 8(6), 10187–10203. |
| [40] | Filus, A., Trzmiel, A., Kuliczkowska-Płaksej, J. Tworowska, U., Jedrzejuk, D., Milewicz, A., Medraś, M. Relationship between vitamin D receptor BsmI and FokI polymorphisms and anthropometric and biochemical parameters describing metabolic syndrome. The aging male: the official journal of the International Society for the Study of the Aging Male, 2008, 11(3), 134–139. |
| [41] | Egan, J. B., Thompson, P. A., Vitanov, M. V., Bartik, L., Jacobs, E. T., Haussler, M. R., Gerner, E. W., Jurutka, P. W. Vitamin D receptor ligands, adenomatous polyposis coli, and the vitamin D receptor FokI polymorphism collectively modulate beta-catenin activity in colon cancer cells. Molecular carcinogenesis, 2010, 49(4), 337–352. |
| [42] | Hustmyer, F. G., DeLuca, H. F., & Peacock, M. ApaI, BsmI, EcoRV and TaqI polymorphisms at the human vitamin D receptor gene locus in Caucasians, blacks and Asians. Human molecular genetics, 1993, 2(4), 487. |
APA Style
Armande, A. A. J., Laurent, S. K., Lydie, B., Stéphane, K. T., Liliane, S. K., et al. (2025). Identification of Genotypes and Allelic Prevalence of the Vitamin D Nuclear Receptor Gene in People Living with Human Immunodeficiency Virus in Côte d'Ivoire. Biochemistry and Molecular Biology, 10(4), 61-70. https://doi.org/10.11648/j.bmb.20251004.11
ACS Style
Armande, A. A. J.; Laurent, S. K.; Lydie, B.; Stéphane, K. T.; Liliane, S. K., et al. Identification of Genotypes and Allelic Prevalence of the Vitamin D Nuclear Receptor Gene in People Living with Human Immunodeficiency Virus in Côte d'Ivoire. Biochem. Mol. Biol. 2025, 10(4), 61-70. doi: 10.11648/j.bmb.20251004.11
AMA Style
Armande AAJ, Laurent SK, Lydie B, Stéphane KT, Liliane SK, et al. Identification of Genotypes and Allelic Prevalence of the Vitamin D Nuclear Receptor Gene in People Living with Human Immunodeficiency Virus in Côte d'Ivoire. Biochem Mol Biol. 2025;10(4):61-70. doi: 10.11648/j.bmb.20251004.11
@article{10.11648/j.bmb.20251004.11,
author = {Aké Aya Jeanne Armande and Séri Kipré Laurent and Boyvin Lydie and Koui Tosséa Stéphane and Siransy Kouabla Liliane and N’guessan Kouassi Raymond and Yapo Adou Francis and Sékongo Yassongui Mamadou and Djaman Allico Joseph},
title = {Identification of Genotypes and Allelic Prevalence of the Vitamin D Nuclear Receptor Gene in People Living with Human Immunodeficiency Virus in Côte d'Ivoire},
journal = {Biochemistry and Molecular Biology},
volume = {10},
number = {4},
pages = {61-70},
doi = {10.11648/j.bmb.20251004.11},
url = {https://doi.org/10.11648/j.bmb.20251004.11},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.bmb.20251004.11},
abstract = {Introduction: Studies conducted in Côte d'Ivoire with HIV patients have revealed disorders in phosphorus/calcium metabolism and 25-hydroxyvitmin D3 status. Due to the important role played by VDR-1,25 (OH)2 D3 in the biological activity of 25-hydroxyvitamin D3, it has become necessary to describe the polymorphism of 4 fragments (Fok-1, Bsm-1, Apa-1, Taq-1) of the nuclear receptor gene Vitamin D3 (VDR) in PLHIV. Methods: The study collected blood samples from 50 individuals, including 32 HIV positive patients and 18 HIV negative as a control. The extraction of genomic DNA from peripheral blood mononuclear cells (PBMC) was done using the QIAamp kit (QIAGEN). After amplification, the PCR products obtained were purified using the "High Pure PCR product purification kit" and sequenced at Eurofilms MWG operon. Results: The prevalence of the Fok-1 mutant C allele was 87.5% (28/32) in PLHIV against 77.8% (14/18) in controls. The single mutant genotype Thr1Met4 was 62.5% in PLHIV versus 55.6% in controls. The prevalence of the mutant allele A (g.63510 A) of Bsm-1 (rs 1544410) was 28% in HIV-infected against 66.7% (12/18) in controls. The mutant GG genotype was more prevalent in 34.4% of PLHIV. Two types of mutant alleles A (g.65023 A) and C (g.65024 C) and the AC mutant genotype of Apa-1 (rs 7975232) were more observed in 28% of those infected with HIV versus 0% in controls. No significant difference was observed between the frequencies of wild T (g.65058 T) allele and mutant C or A (g.65058 C or A) of Taq-1 (rs 731236) (p = 0.47). The wild genotype Ile352 was the most common regarding the Taq-1 polymorphism. Ten PLHIV and controls have the combination Fok-1 + Bsm-1 in common. The combinations Fok-1 + Bsm-1 + Apa-1 and Fok-1 + Apa-1 were specific to PLHIV. Conclusion: The blood sample of PLHIV subjects is characterized by polymorphisms affecting Fok-1, Bsm-1, and Apa-1. In addition, the combination of Fok-1 and Bsm-1 was observed in both PLHIV patients and controls.},
year = {2025}
}
TY - JOUR T1 - Identification of Genotypes and Allelic Prevalence of the Vitamin D Nuclear Receptor Gene in People Living with Human Immunodeficiency Virus in Côte d'Ivoire AU - Aké Aya Jeanne Armande AU - Séri Kipré Laurent AU - Boyvin Lydie AU - Koui Tosséa Stéphane AU - Siransy Kouabla Liliane AU - N’guessan Kouassi Raymond AU - Yapo Adou Francis AU - Sékongo Yassongui Mamadou AU - Djaman Allico Joseph Y1 - 2025/10/09 PY - 2025 N1 - https://doi.org/10.11648/j.bmb.20251004.11 DO - 10.11648/j.bmb.20251004.11 T2 - Biochemistry and Molecular Biology JF - Biochemistry and Molecular Biology JO - Biochemistry and Molecular Biology SP - 61 EP - 70 PB - Science Publishing Group SN - 2575-5048 UR - https://doi.org/10.11648/j.bmb.20251004.11 AB - Introduction: Studies conducted in Côte d'Ivoire with HIV patients have revealed disorders in phosphorus/calcium metabolism and 25-hydroxyvitmin D3 status. Due to the important role played by VDR-1,25 (OH)2 D3 in the biological activity of 25-hydroxyvitamin D3, it has become necessary to describe the polymorphism of 4 fragments (Fok-1, Bsm-1, Apa-1, Taq-1) of the nuclear receptor gene Vitamin D3 (VDR) in PLHIV. Methods: The study collected blood samples from 50 individuals, including 32 HIV positive patients and 18 HIV negative as a control. The extraction of genomic DNA from peripheral blood mononuclear cells (PBMC) was done using the QIAamp kit (QIAGEN). After amplification, the PCR products obtained were purified using the "High Pure PCR product purification kit" and sequenced at Eurofilms MWG operon. Results: The prevalence of the Fok-1 mutant C allele was 87.5% (28/32) in PLHIV against 77.8% (14/18) in controls. The single mutant genotype Thr1Met4 was 62.5% in PLHIV versus 55.6% in controls. The prevalence of the mutant allele A (g.63510 A) of Bsm-1 (rs 1544410) was 28% in HIV-infected against 66.7% (12/18) in controls. The mutant GG genotype was more prevalent in 34.4% of PLHIV. Two types of mutant alleles A (g.65023 A) and C (g.65024 C) and the AC mutant genotype of Apa-1 (rs 7975232) were more observed in 28% of those infected with HIV versus 0% in controls. No significant difference was observed between the frequencies of wild T (g.65058 T) allele and mutant C or A (g.65058 C or A) of Taq-1 (rs 731236) (p = 0.47). The wild genotype Ile352 was the most common regarding the Taq-1 polymorphism. Ten PLHIV and controls have the combination Fok-1 + Bsm-1 in common. The combinations Fok-1 + Bsm-1 + Apa-1 and Fok-1 + Apa-1 were specific to PLHIV. Conclusion: The blood sample of PLHIV subjects is characterized by polymorphisms affecting Fok-1, Bsm-1, and Apa-1. In addition, the combination of Fok-1 and Bsm-1 was observed in both PLHIV patients and controls. VL - 10 IS - 4 ER -
Quality Control Laboratory, National Blood Transfusion Center of Côte d'Ivoire (CNTSCI), Abidjan, Côte d'Ivoire
Research Fields: Biochemistry, Molecular Biology, Immunology, Hematology, Infectiology, Immunogenetics
Department of Medical and Fundamental Biochemistry, Institut Pasteur of Côte d'Ivoire (IPCI), Abidjan, Côte d'Ivoire
Research Fields: Biochemistry, Functional and molecular Biology, Functional nutrition, Toxicology, Pharmacology, Proteomics, Metabolomics
Department of Medical and Fundamental Biochemistry, Institut Pasteur of Côte d'Ivoire (IPCI), Abidjan, Côte d'Ivoire
Research Fields: Biochemistry, Molecular Biology, Microbiology, hematology, Pharmacology
Quality Control Laboratory, National Blood Transfusion Center of Côte d'Ivoire (CNTSCI), Abidjan, Côte d'Ivoire
Research Fields: Genetic, Molecular Biology, Biology, Immunogenetics
Quality Control Laboratory, National Blood Transfusion Center of Côte d'Ivoire (CNTSCI), Abidjan, Côte d'Ivoire; Allergy Immunology Laboratory, Félix Houphouët-Boigny University, Abidjan, Côte d'Ivoire
Research Fields: Immunology, Allergology, Biochemistry, Molecular Biology, Histocompatibility, Immunogenetics
Department of Tuberculosis, Institut Pasteur of Côte d'Ivoire (IPCI), Abidjan, Côte d'Ivoire
Research Fields: Molecular Biology, Microbiology, Tuberculosis
Biology and Health Laboratory, Félix Houphouët-Boigny University, Abidjan, Côte d'Ivoire
Research Fields: Biochemistry, Hematology, Pharmacology
Quality Control Laboratory, National Blood Transfusion Center of Côte d'Ivoire (CNTSCI), Abidjan, Côte d'Ivoire
Research Fields: Hematology, Immunology, Blood transfusion, Hemoglobinopathies
Department of Medical and Fundamental Biochemistry, Institut Pasteur of Côte d'Ivoire (IPCI), Abidjan, Côte d'Ivoire; Biology and Health Laboratory, Félix Houphouët-Boigny University, Abidjan, Côte d'Ivoire
Research Fields: Biochemistry, Parasitology, Molecular Biology, Microbiology, Hematology, Pharmacology
Information