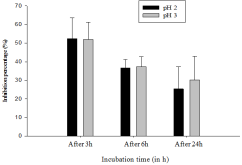
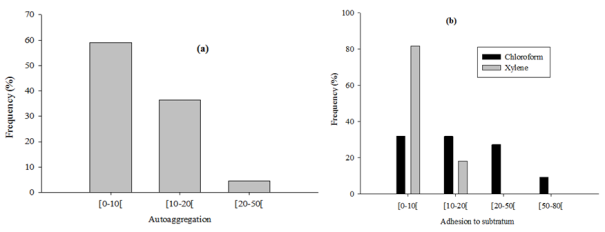
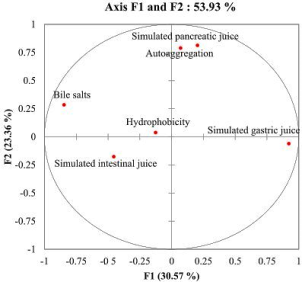
The microorganisms intended for use as probiotics in food formulation should exert health benefit effects and be regarded as safe for animals and humans uses. The aim of this study was to evaluate the probiotic potential of lactic acid bacteria (LAB) isolated from pendidam and kindirmou, two traditional fermented milks (TFM) produced in the Adamawa region (Cameroon). Twenty-five samples (pendidam: 13 and kindirmou: 12) were randomly collected in five markets of Ngaoundere (n = 17 samples) and Meiganga (n = 8 samples). These samples were screened for their antimicrobial activity, and nine TFMs were retained. Lactic acid bacteria were isolated from these samples and their antimicrobial activity was already evaluated. Based on the inhibition zone, twenty-two LABs were retained and examined in vitro for potential probiotic properties based on their low pH tolerance, resistance to bile salts, tolerance to simulated gastrointestinal juices, hydrophobicity, autoaggregation, gelatinase and hemolytic activities. The outcome of these parameters studied was used as input data for a principal component analysis (PCA) to select the most promising isolate, and the six potential probiotic isolates were characterized through a biochemical profile. The characterized isolates have been identified as Lactiplantibacillus plantarum, Lacticaseibacillus casei, and Lactococcus lactis. Traditional fermented milks contain LAB with important properties that can be utilized in the formulation of functional foods.
| Published in | Advances in Biochemistry (Volume 12, Issue 2) |
| DOI | 10.11648/j.ab.20241202.11 |
| Page(s) | 35-48 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2024. Published by Science Publishing Group |
Probiotic, Lactic Acid Bacteria, Pendidam, Kindirmou, Cameroon
LAB | Diameter of inhibition (mm) | ||||
|---|---|---|---|---|---|
M. morganii | P. mirabilis | L. monocytogenes | B. cereus | K. pneumoniae | |
PD1 | 48.0 ± 1.4k | 41.2 ± 0.3i | 65.5 ± 0.4jk | 57.5 ± 0.7hi | 50.5 ± 0.7m |
PD2 | 48.5 ± 0.7k | 34.3 ± 0.4h | 72.5 ± 0.6m | 64.6 ± 0.8e | 25.0 ± 0.6b |
PD3 | 32.0 ± 1.4fg | 31.8 ± 0.2fgh | 51.0 ± 1.0c | 45.5 ± 0.5d | 44.4 ± 0.5k |
PD4 | 35.5 ± 0.7i | 21.5 ± 0.3bc | 66.5 ± 0.6kl | 59.7 ± 0.3jk | 19.6 ± 0.8a |
PD5 | 24.5 ± 0.3b | 14.5 ± 0.7a | 53.5 ± 0.4d | 57.5 ± 0.6hi | 30.2 ± 0.3e |
PD6 | 33.7 ± 0.3ghi | 39.7 ± 1.0i | 70.5 ± 0.7m | 60.4 ± 0.5k | 34.4 ± 0.5h |
PD8 | 29.0 ± 0.2e | 29.6 ± 0.8def | 56.0 ± 0.8e | 57.5 ± 0.7hi | 30.5 ± 0.7ef |
PB1 | 31.7 ± 0.4fg | 21.3 ± 0.4bc | 57.0 ± 1.0ef | 52.7 ± 0.5f | 31.6 ± 0.4fg |
PB2 | 24.2 ± 0.3b | 32.4 ± 0.5gh | 57.5 ± 0.5ef | 56.5 ± 0.4d | 28.2 ± 0.2d |
PB3 | 32.0 ± 1.2bc | 41.7 ± 0.8i | 63.4 ± 0.7hij | 57.5 ± 1.0hi | 34.5 ± 0.7h |
PB4 | 34.0 ± 0.8ghi | 31.0 ± 1.0efg | 64.5 ± 0.7ijk | 54.6 ± 0.2h | 39.3 ± 0.4i |
PD9 | 21.0 ± 1.0a | 34.0 ± 0.6h | 38.7 ± 0.3b | 30.7 ± 0.8a | 24.5 ± 0.2b |
PD10 | 35.0 ± 0.7hi | 27.1 ± 1.0d | 61.2 ± 1.0gh | 54.6 ± 0.6g | 23.7 ± 0.3b |
PD11 | 32.7 ± 1.0fgh | 29.5 ± 0.7def | 62.5 ± 0.8ghi | 58.4 ± 0.5ij | 26.4 ± 0.5c |
PD12 | 28.4 ± 0.5de | 27.2 ± 1.2d | 56.0 ± 0.6e | 40.5 ± 0.3c | 34.5 ± 0.2h |
PG1 | 26.6 ± 0.8cd | 22.2 ± 0.3bc | 61.7 ± 0.3gh | 58.4 ± 0.5ij | 26.5 ± 0.8c |
PG2 | 24.7 ± 0.8bc | 20.0 ± 1.0b | 58.5 ± 0.6f | 50.6 ± 0.8 | 34.3 ± 0.4h |
PG3 | 39.6 ± 0.8j | 31.0 ± 0.6efg | 67.7 ± 0.2l | 61.2 ± 0.2k | 42.1 ± 0.2j |
PG4 | 33.9 ± 0.1ghi | 29.5 ± 0.7def | 68.0 ± 0.2l | 58.2 ± 0.3ij | 44.1 ± 0.2k |
PG5 | 39.6 ± 0.5j | 30.0 ± 1.0efg | 27.0 ± 1.0a | 34.5 ± 0.7b | 32.3 ± 0.4g |
KM1 | 30.5 ± 0.7ef | 28.7 ± 1.0de | 60.8 ± 1.0g | 57.3 ± 0.4hi | 24.3 ± 0.3b |
KB3 | 31.2 ± 1.0f | 23.5 ± 0.7c | 62.2 ± 0.3gh | 56.4 ± 0.5h | 48.2 ± 0.4l |
LAB | Diameter of inhibition (mm) | |||||
|---|---|---|---|---|---|---|
C. parapsilosis | S. cerevisiae | C. albicans | R. mucilaginosa | D. hansenii | K. marxianus | |
PD1 | 16.5 ± 0.7a | 0a | 16.2 ± 0.3b | 14.5 ± 0.7b | 26.1 ± 0.1i | 26.2 ± 0.4k |
PD2 | 30.3 ± 0.2h | 0a | 24.4 ± 0.5e | 18.4 ± 0.6c | 20.5 ± 0.7g | 14.5 ± 0.7c |
PD3 | 32.5 ± 0.6i | 0a | 29.5 ± 0.7hi | 14.6 ± 0.8b | 18.1 ± 0.1ef | 20.2 ± 0.4fg |
PD4 | 22.4 ± 0.5e | 0a | 24.4 ± 0.6e | 18.4 ± 0.6c | 14.2 ± 0.3d | 22.7 ± 1.0k |
PD5 | 16.1 ± 0.1a | 0a | 26.3 ± 0.4f | 22.1 ± 0.1f | 10.5 ± 0.7c | 12.5 ± 0.2b |
PD6 | 21.5 ± 0.5cde | 0a | 14.8 ± 0.1b | 14.4 ± 0.6b | 24.0 ± 0.0h | 18.5 ± 0.7e |
PD8 | 25.0 ± 0.4f | 0a | 24.6 ± 0.8e | 22.5 ± 0.7f | 8.4 ± 0.6b | 21.5 ± 0.3ghi |
PB1 | 22.2 ± 0.8de | 0a | 20.4 ± 0.5c | 10.4 ± 0.6a | 8.5 ± 0.7b | 18.3 ± 0.4e |
PB2 | 16.4 ± 0.6a | 0a | 24.3 ± 0.4e | 20.3 ± 0.4d | 26.2 ± 0.4i | 21.8 ± 0.3hi |
PB3 | 20.2 ± 0.2bc | 0a | 34.5 ± 0.7j | 21.7 ± 0.4ef | 17.6 ± 3.3ef | 12.5 ± 0.7b |
PB4 | 20.5 ± 0.2bc | 0a | 36.2 ± 0.3i | 28.1 ± 0.1i | 16.2 ± 0.3e | 10.7 ± 1.1a |
PD9 | 30.4 ± 0.4h | 0a | 30.4 ± 0.5k | 10.3 ± 0.5a | 19.0 ± 0.0fg | 16.2 ± 0.4d |
PD10 | 28.5 ± 0.3g | 0a | 26.6 ± 0.8f | 14.2 ± 0.4b | 6.5 ± 0.7a | 12.5 ± 0.7b |
PD11 | 28.2 ± 0.5g | 0a | 22.3 ± 0.4d | 20.8 ± 0.3de | 20.2 ± 0.3g | 18.4 ± 0.6e |
PD12 | 29.4 ± 0.1gh | 0a | 39.5 ± 0.7l | 24.2 ± 0.3g | 22.5 ± 0.7h | 21.0 ± 1.4gh |
PG1 | 26.1 ± 1.0f | 11.0 ± 0.8e | 26.8 ± 1.0fg | 25.3 ± 0.4h | 17.6 ± 0.8ef | 26.5 ± 0.6k |
PG2 | 28.8 ± 0.8gh | 9.2 ± 0.4d | 24.5 ± 0.7e | 18.2 ± 0.4c | 23.2 ± 0.3h | 19.3 ± 0.4ef |
PG3 | 20.6 ± 0.6bcd | 3.4 ± 0.5b | 34.4 ± 0.5j | 22.1 ± 0.1f | 20.3 ± 0.4g | 20.2 ± 0.3fg |
PG4 | 22.2 ± 0.3de | 6.7 ± 0.1c | 34.5 ± 0.7j | 18.3 ± 0.4c | 23.4 ± 0.6h | 24.4 ± 0.6k |
PG5 | 19.5 ± 0.7b | 0a | 10.9 ± 0.2a | 24.2 ± 0.3g | 16.2 ± 0.4 | 19.2 ± 0.4ef |
KM1 | 23.0 ± 1.0e | 0a | 20.2 ± 0.6c | 28.4 ± 0.6i | 20.5 ± 0.7g | 26.2 ± 0.3k |
KB3 | 22.7 ± 1.0e | 0a | 28.2 ± 0.1gh | 24.3 ± 0.4g | 29.6 ± 0.8j | 30.5 ± 0.7l |
Isolates | Initial count | Bile salts (%) | SGJ | SPJ | SIJ | |
|---|---|---|---|---|---|---|
0.2 | 0.45 | 180 min | 90 min | 240 min | ||
KB3 | 7.76 | 6.75 ± 0.21ef | 6.21 ± 0.11efg | 7.59 ± 0.20abc | 7.61 ± 0.34bcde | 7.67 ± 0.02fg |
KM1 | 7.66 | 6.18 ± 0.30d | 5.86 ± 0.15de | 7.57 ± 0.09abc | 7.44 ± 0.22bc | 7.50 ± 0.20efg |
PB1 | 7.68 | 6.27 ± 0.07de | 5.99 ± 0.21ef | 7.76 ± 0.12bcde | 9.05 ± 0.41f | 7.38 ± 0.04ef |
PB2 | 9.11 | 8.21 ± 0.26h | 7.21 ± 0.10jkl | 9.26 ± 0.08fgh | 8.79 ± 0.19f | 8.9 ± 0.01ij |
PB3 | 7.77 | 6.25 ± 0.15de | 6.12 ± 0.13ef | 7.79 ± 0.02bcde | 7.49 ± 0.26bcd | 5.57 ± 0.22c |
PB4 | 7.79 | 4.75 ± 0.21b | 4.83 ± 0.11b | 7.81 ± 0.14bcde | 7.88 ± 0.24cde | 5.61 ± 0.19c |
PD1 | 9.11 | 5.38 ± 0.02c | 5.39 ± 0.10c | 9.38 ± 0.12gh | 8.84 ± 0.13f | 4.44 ± 0.07b |
PD10 | 7.91 | 7.52 ± 0.23g | 6.88 ± 0.22ij | 7.72 ± 0.07abcd | 8.07 ± 0.23e | 6.60 ± 0.09d |
PD11 | 7.72 | 7.33 ± 0.42g | 6.52 ± 0.05ghi | 7.54 ± 0.07ab | 7.57 ± 0.11bcde | 7.35 ± 0.09e |
PD12 | 7.65 | 5.47 ± 0.48c | 4.85 ± 0.16b | 8.09 ± 0.09de | 7.22 ± 0.04ab | 6.47 ± 0.12d |
PD2 | 9.18 | 7.48 ± 0.05g | 7.25 ± 0.12kl | 8.85 ± 0.31f | 9.8 ± 0.35h | 9.14 ± 0.21j |
PD3 | 9.08 | 3.25 ± 0.23a | 2.70 ± 0.14a | 9.43 ± 0.24h | 8.91 ± 0.27f | 8.64 ± 0.08i |
PD4 | 7.89 | 6.24 ± 0.23de | 5.64 ± 0.05cd | 8.14 ± 0.13e | 8.87 ± 0.27f | 3.50 ± 0.13a |
PD5 | 9.11 | 8.07 ± 0.19h | 7.56 ± 0.07lm | 9.00 ± 0.22fg | 9.19 ± 0.24fg | 8.09 ± 0.01h |
PD6 | 9.15 | 7.84 ± 0.08gh | 7.55 ± 0.19lm | 9.19 ± 0.43fgh | 9.61 ± 0.12gh | 8.71 ± 0.14i |
PD8 | 7.83 | 6.73 ± 0.03ef | 6.67 ± 0.16i | 7.99 ± 0.14cde | 8.03 ± 0.43de | 7.68 ± 0.04g |
PD9 | 9.03 | 7.47 ± 0.36g | 6.63 ± 0.06hi | 9.19 ± 0.22fgh | 9.06 ± 0.29f | 8.74 ± 0.16i |
PG1 | 7.96 | 7.48 ± 0.07g | 7.12 ± 0.16jk | 7.95 ± 0.01bcde | 6.85 ± 0.23a | 7.51 ± 0.03efg |
PG2 | 9.04 | 8.29 ± 0.08h | 8.23 ± 0.12n | 8.94 ± 0.11f | 8.92 ± 0.21f | 8.88 ± 0.14ij |
PG3 | 7.69 | 6.57 ± 0.16def | 6.30 ± 0.21fgh | 7.64 ± 0.14abc | 7.56 ± 0.02bcde | 7.58 ± 0.09efg |
PG4 | 7.45 | 6.84 ± 0.19f | 6.26 ± 0.21fg | 7.33 ± 0.06a | 7.81 ± 0.08cde | 7.33 ± 0.02e |
PG5 | 9.03 | 8.18 ± 0.01h | 7.76 ± 0.31m | 8.85 ± 0.13f | 8.78 ± 0.04f | 8.83 ± 0.18i |
Variables | F1 | F2 | F3 |
|---|---|---|---|
Hydrophobicity | -0.126 | 0.038 | 0.795 |
Autoaggregation | 0.07 | 0.79 | -0.235 |
Simulated gastric juice | 0.922 | -0.061 | -0.149 |
Simulated pancreatic juice | 0.203 | 0.813 | 0.026 |
Simulated intestinal juice | -0.455 | -0.176 | -0.635 |
Bile salts | -0.846 | 0.284 | 0.047 |
| [1] | FAO/WHO, “FAO/WHO expert consultation on evaluation of health and nutritional properties of probiotics in food including powder milk with live lactic acid bacteria,” 2002. |
| [2] | K. Angmo, A. Kumari, Savitri, and T. C. Bhalla, “Probiotic characterization of lactic acid bacteria isolated from fermented foods and beverage of Ladakh,” LWT - Food Sci. Technol., vol. 66, no. October, pp. 428–435, 2016, |
| [3] | A. Abushelaibi, S. Al-Mahadin, K. El-Tarabily, N. P. Shah, and M. Ayyash, “Characterization of potential probiotic lactic acid bacteria isolated from camel milk,” LWT - Food Sci. Technol., vol. 79, pp. 316–325, 2017, |
| [4] | Z. Guo, J. Wang, L. Yan, W. Chen, X. ming Liu, and H. ping Zhang, “In vitro comparison of probiotic properties of Lactobacillus casei Zhang, a potential new probiotic, with selected probiotic strains,” LWT - Food Sci. Technol., vol. 42, no. 10, pp. 1640–1646, 2009, |
| [5] | Y. Moreno, M. C. Collado, M. A. Ferrús, J. M. Cobo, E. Hernández, and M. Hernández, “Viability assessment of lactic acid bacteria in commercial dairy products stored at 4°C using LIVE/DEAD® BacLightTM staining and conventional plate counts,” Int. J. Food Sci. Technol., vol. 41, no. 3, pp. 275–280, 2006, |
| [6] | T. Faye, A. Tamburello, G. E. Vegarud, and S. Skeie, “Survival of lactic acid bacteria from fermented milks in an in vitro digestion model exploiting sequential incubation in human gastric and duodenum juice,” J. Dairy Sci., vol. 95, no. 2, pp. 558–566, 2012, |
| [7] | F. M. de Sant’Anna et al., “Assessment of the probiotic potential of lactic acid bacteria isolated from Minas artisanal cheese produced in the Campo das Vertentes region, Brazil,” Int. J. Dairy Technol., vol. 70, no. 4, pp. 592–601, 2017, |
| [8] | L. De Vuyst, M. R. Foulquié Moreno, and H. Revets, “Screening for enterocins and detection of hemolysin and vancomycin resistance in enterococci of different origins,” Int. J. Food Microbiol., vol. 84, no. 3, pp. 299–318, 2003, |
| [9] | M. Sidira et al., “Effect of probiotic-Fermented milk administration on gastrointestinal survival of Lactobacillus casei ATCC 393 and modulation of intestinal microbial flora,” J. Mol. Microbiol. Biotechnol., vol. 19, no. 4, pp. 224–230, 2011, |
| [10] | D. V. Sieladie, F. N. Zambou, P. M. Kaktcham, A. Cresci, and F. Fonteh, “Probiotic properties of lactobacilli strains isolated from raw cow milk in the western highlands of Cameroon,” Innov. Rom. Food Biotechnol., vol. 9, no. August 2015, pp. 12–28, 2011. |
| [11] | M. P. Vélez, K. Hermans, T. L. A. Verhoeven, S. E. Lebeer, J. Vanderleyden, and S. C. J. De Keersmaecker, “Identification and characterization of starter lactic acid bacteria and probiotics from Columbian dairy products,” J. Appl. Microbiol., vol. 103, no. 3, pp. 666–674, 2007, |
| [12] | M. Iranmanesh, H. Ezzatpanah, and N. Mojgani, “Antibacterial activity and cholesterol assimilation of lactic acid bacteria isolated from traditional Iranian dairy products,” LWT - Food Sci. Technol., vol. 58, no. 2, pp. 355–359, 2014, |
| [13] | J. M. Mathara et al., “Functional characteristics of Lactobacillus spp. from traditional Maasai fermented milk products in Kenya,” Int. J. Food Microbiol., vol. 126, no. 1–2, pp. 57–64, 2008, |
| [14] | M. Abdelbasset and K. Djamila, “Antimicrobial activity of autochthonous lactic acid bacteria isolated from Algerian traditional fermented milk Raib,” African J. Biotechnol., vol. 7, no. 16, pp. 2908–2914, 2008, |
| [15] | D. Guilhem, “Le lait de vache dans les sociétés peules Pratiques alimentaires et symbolisme d ’ un critère identitaire,” pp. 1–8, 2006. |
| [16] | N. M. M. Tchamba et al., “Assessment of Probiotic Potential of Lactic Acid Bacteria Isolated from Bottle Gourds (Calabash) of Milk Fermentation of Mbéré, Cameroon,” J. Pharm. Res. Int., vol. 33, pp. 356–369, 2021, |
| [17] | W. Liu et al., “Isolation and identification of lactic acid bacteria from Tarag in Eastern Inner Mongolia of China by 16S rRNA sequences and DGGE analysis,” Microbiol. Res., vol. 167, no. 2, pp. 110–115, 2012, |
| [18] | D. Libouga, J. Essia Ngang, and H. Halilou, “Qualité de quelques laits fermentés camerounais,” Sci. Aliments, vol. 25, no. 1, pp. 53–66, Feb. 2005, |
| [19] | J. Maïworé, L. Tatsadjieu, and I. Piro-metayer, “Identification of yeasts present in artisanal yoghurt and traditionally fermented milks consumed in the northern part of Cameroon,” vol. 6, pp. 0–8, 2019, |
| [20] | A. Mbawala, M. M. Pahane, and H. T. Mouafo, “Effect of manufacturing practices on the microbiological quality of fermented milk (Pendidam) of some localities of Ngaoundere (Cameroon),” Int. J. Curr. Microbiol. Appl. Sci., vol. 3, no. 11, pp. 71–81, 2014. |
| [21] | T. H. Mouafo, A. Mbawala, and R. Ndjouenkeu, “Effect of different carbon sources on biosurfactants’ production by three strains of Lactobacillus spp.,” Biomed Res. Int., vol. 2018, 2018, |
| [22] | M. T. Hippolyte, M. Augustin, T. M. Hervé, N. Robert, and S. Devappa, “Application of response surface methodology to improve the production of antimicrobial biosurfactants by Lactobacillus paracasei subsp. tolerans N2 using sugar cane molasses as substrate,” Bioresour. Bioprocess., vol. 5, no. 1, 2018, |
| [23] | Codex Alimentarus, “Méthodes d’analyse et d’échantillonnage recommandées, Première partie, Méthodes d’analyse et d’échantillonnage par ordre alphabétique des catégories et noms de produit,” pp. 18–24, 1999. |
| [24] | A. Mbawala, P. Y. Mahbou, H. T. Mouafo, and L. N. Tatsadjieu, “Antibacterial activity of some lactic acid bacteria isolated from a local fermented milk product (Pendidam) in Ngaoundere, Cameroon,” J. Anim. Plant Sci., vol. 23, no. 1, pp. 157–166, 2013. |
| [25] | J. C. de Man, M. Rogosa, and M. E. Sharpe, “A medium used for the cultivation of Lactobacilli,” J Appl Bacteriol, vol. 23, pp. 130–135, 1960. |
| [26] | H. P. Fleming, J. L. Etchells, and R. N. Costilow, “Microbial inhibition by an isolate of Pediococcus from cucumber brines.,” Appl. Microbiol., vol. 30, no. 6, pp. 1040–2, 1975. |
| [27] | B. Kos, J. Šušković, S. Vuković, M. Sǐmpraga, J. Frece, and S. Matošić, “Adhesion and aggregation ability of probiotic strain Lactobacillus acidophilus M92,” J. Appl. Microbiol., vol. 94, no. 6, pp. 981–987, 2003, |
| [28] | CASFM/EUCAST, “Comité de l’Antibiogramme de la Société Française de Microbiologie,” Eur. Soc. Clin. Microbiol. Infect. Dis., vol. V1.0 2019, pp. S3–S4, 2019, |
| [29] | T. J. Eaton and M. J. Gasson, “Molecular Screening of Enterococcus Virulence Determinants and Potential for Genetic Exchange between Food and Medical Isolates” Appl. Environ. Microbiol., vol. 67, no. 4, pp. 1628–1635, 2001, |
| [30] | C. Jiwoua and J. B. Millière, “Flore lactique et entérocoques du lait caillé (Pindidam) produit dans l’Adamaoua (Cameroun),” Lait, vol. 70, no. 5–6, pp. 475–486, 1990. |
| [31] | K. Voulgari, M. Hatzikamari, A. Delepoglou, P. Georgakopoulos, E. Litopoulou-Tzanetaki, and N. Tzanetakis, “Antifungal activity of non-starter lactic acid bacteria isolates from dairy products,” Food Control, vol. 21, no. 2, pp. 136–142, 2010, |
| [32] | H. J. Song and J. Richard, “Antilisterial activity of three bacteriocins used at sub minimal inhibitory concentrations and cross-resistance of the survivors,” Int. J. Food Microbiol., vol. 36, no. 2–3, pp. 155–161, 1997, |
| [33] | J. Magnusson, K. Ström, S. Roos, J. Sjögren, and J. Schnürer, “Broad and complex antifungal activity among environmental isolates of lactic acid bacteria,” FEMS Microbiol. Lett., vol. 219, no. 1, pp. 129–135, 2003, |
| [34] | J. Magnusson and J. Schnürer, “Lactobacillus coryniformis subsp. coryniformis Strain Si3 Produces a Broad-Spectrum Proteinaceous Antifungal Compound,” Appl. Environ. Microbiol., vol. 67, no. 1, pp. 1–5, 2001, |
| [35] | J. Schnürer and J. Magnusson, “Antifungal lactic acid bacteria as biopreservatives,” Trends Food Sci. Technol., vol. 16, no. 1–3, pp. 70–78, 2005, |
| [36] | D. K. D. Dalié, A. M. Deschamps, and F. Richard-Forget, “Lactic acid bacteria - Potential for control of mould growth and mycotoxins: A review,” Food Control, vol. 21, no. 4, pp. 370–380, 2010, |
| [37] | A. Mechai, M. Debabza, and D. Kirane, “Screening of technological and probiotic properties of lactic acid bacteria isolated from Algerian traditional fermented milk products,” Int. Food Res. J., vol. 21, no. 6, pp. 2451–2457, 2014. |
| [38] | T. P. Singh, G. Kaur, R. K. Malik, U. Schillinger, C. Guigas, and S. Kapila, “Characterization of Intestinal Lactobacillus reuteri Strains as Potential Probiotics,” Probiotics Antimicrob. Proteins, vol. 4, no. 1, pp. 47–58, Mar. 2012, |
| [39] | M. Guetouache and B. Guessas, “Characterization and identification of lactic acid bacteria isolated from traditional cheese (Klila) prepared from cows milk,” African J. Microbiol. Res., vol. 9, no. 2, pp. 71–77, 2015, |
| [40] | C. M. Kalui, J. M. Mathara, P. M. Kutima, C. Kiiyukia, and L. E. Wongo, “Functional characteristics of Lactobacillus plantarum and Lactobacillus rhamnosus from ikii, a Kenyan traditional fermented maize porridge,” African J. Biotechnol., vol. 8, no. 18, pp. 4363–4373, 2009, |
| [41] | M. T. Liong and N. P. Shah, “Acid and bile tolerance and cholesterol removal ability of lactobacilli strains,” J. Dairy Sci., vol. 88, no. 1, pp. 55–66, 2005, |
| [42] | P. D. Cotter and C. Hill, “Surviving the Acid Test: Responses of Gram-Positive Bacteria to Low pH,” Microbiol. Mol. Biol. Rev., vol. 67, no. 3, pp. 429–453, 2003, |
| [43] | W. P. Charteris, P. M. Kelly, L. Morelli, and J. K. Collins, “Antibiotic susceptibility of potentially probiotic Bifidobacterium isolates from the human gastrointestinal tract,” Lett. Appl. Microbiol., vol. 26, no. 5, pp. 333–337, 1998, |
| [44] | P. A. Maragkoudakis, G. Zoumpopoulou, C. Miaris, G. Kalantzopoulos, B. Pot, and E. Tsakalidou, “Probiotic potential of Lactobacillus strains isolated from dairy products,” Int. Dairy J., vol. 16, no. 3, pp. 189–199, 2006, |
| [45] | B. E. Minelli et al., “Assessment of novel probiotic Lactobacillus casei strains for the production of functional dairy foods,” Int. Dairy J., vol. 14, no. 8, pp. 723–736, 2004, |
| [46] | S. Salminen et al., “Demonstration of safety of probiotics - A review,” Int. J. Food Microbiol., vol. 44, no. 1–2, pp. 93–106, 1998, |
| [47] | D. Donohue and S. Salminen, “Safety of probiotic bacteria,” Asian Pacific J. Clin. Nutr., vol. 5, pp. 25–28, 1996. |
| [48] | E. Muñoz-Atienza et al., “Antimicrobial activity, antibiotic susceptibility and virulence factors of Lactic Acid Bacteria of aquatic origin intended for use as probiotics in aquaculture,” BMC Microbiol., vol. 13, no. 1, 2013, |
| [49] | D. J. Birri, D. A. Brede, G. T. Tessema, and I. F. Nes, “Bacteriocin Production, Antibiotic Susceptibility and Prevalence of Haemolytic and Gelatinase Activity in Faecal Lactic Acid Bacteria Isolated from Healthy Ethiopian Infants,” Microb. Ecol., vol. 65, no. 2, pp. 504–516, 2013, |
| [50] | A. Hummel, W. H. Holzapfel, and C. M. A. P. Franz, “Characterisation and transfer of antibiotic resistance genes from enterococci isolated from food,” Syst. Appl. Microbiol., vol. 30, no. 1, pp. 1–7, 2007, |
| [51] | A. S. Hummel, C. Hertel, W. H. Holzapfel, and C. M. A. P. Franz, “Antibiotic resistances of starter and probiotic strains of lactic acid bacteria,” Appl. Environ. Microbiol., vol. 73, no. 3, pp. 730–739, 2007, |
| [52] | M. S. Ammor, A. Belén Flórez, and B. Mayo, “Antibiotic resistance in non-enterococcal lactic acid bacteria and bifidobacteria,” Food Microbiol., vol. 24, no. 6, pp. 559–570, 2007, |
| [53] | M. C. Collado, J. Meriluoto, and S. Salminen, “Adhesion and aggregation properties of probiotic and pathogen strains,” Eur. Food Res. Technol., vol. 226, no. 5, pp. 1065–1073, 2008, |
| [54] | Y. Bao et al., “Screening of potential probiotic properties of Lactobacillus fermentum isolated from traditional dairy products,” Food Control, vol. 21, no. 5, pp. 695–701, 2010, |
| [55] | T. García-Cayuela et al., “Adhesion abilities of dairy Lactobacillus plantarum strains showing an aggregation phenotype,” Food Res. Int., vol. 57, pp. 44–50, 2014, |
| [56] | T. Janković, J. Frece, M. Abram, and I. Gobin, “Aggregation ability of potential probiotic Lactobacillus plantarum strains,” Int. J. Sanit. Eng. Res., vol. 6, no. 1, pp. 19–24, 2012. |
| [57] | T. Wadstroum, K. Andersson, M. Sydow, L. Axelsson, S. Lindgren, and B. Gullmar, “Surface properties of lactobacilli isolated from the small intestine of pigs,” J. Appl. Bacteriol., vol. 62, no. 6, pp. 513–520, 1987, |
| [58] | Q. Li, X. Liu, and J. Zhou, “Aggregation and adhesion abilities of 18 lactic acid bacteria strains isolated from traditional fermented food,” Int. J. Agric. Policy Res., vol. 3, no. February, pp. 84–92, 2015. |
| [59] | J. D. Greene and T. R. Klaenhammer, “Factors involved in adherence of lactobacilli to human Caco-2 cells,” Appl. Environ. Microbiol., vol. 60, no. 12, pp. 4487–4494, 1994. |
| [60] | E. M. Tuomola, A. C. Ouwehand, and S. J. Salminen, “Chemical, physical and enzymatic pre-treatments of probiotic lactobacilli alter their adhesion to human intestinal mucus glycoproteins,” Int. J. Food Microbiol., vol. 60, no. 1, pp. 75–81, 2000, |
| [61] | C. Pelletier, C. Bouley, C. Cayuela, S. Bouttier, P. Bourlioux, and M. N. Bellon-Fontaine, “Cell surface characteristics of Lactobacillus casei subsp. casei, Lactobacillus paracasei subsp. paracasei, and Lactobacillus rhamnosus strains,” Appl. Environ. Microbiol., vol. 63, no. 5, pp. 1725–1731, 1997. |
| [62] | D. Granato et al., “Cell surface-associated lipoteichoic acid acts as an adhesion factor for attachment of Lactobacillus johnsonii La1 to human enterocyte-like Caco-2 cells,” Appl. Environ. Microbiol., vol. 65, no. 3, pp. 1071–1077, 1999. |
| [63] | J. Frece, B. Kos, I. K. Svetec, Z. Zgaga, V. Mrša, and J. Šušković, “Importance of S-layer proteins in probiotic activity of Lactobacillus acidophilus M92,” J. Appl. Microbiol., vol. 98, no. 2, pp. 285–292, 2005, |
| [64] | M. L. Sorongon, R. A. Bloodgood, and R. P. Burchard, “Hydrophobicity, Adhesion, and Surface-Exposed Proteins of Gliding Bacteria,” Appl. Environ. Microbiol., vol. 57, no. 11, pp. 3193–3199, 1991. |
APA Style
Steve, N. S. F., Adji, M. B., Alphonse, S. T., Augustin, M., Leopold, T. N. (2024). Isolation and Probiotic Characterization of Lactic Acid Bacteria from Kindirmou and Pendidam in Adamawa Region (Cameroon). Advances in Biochemistry, 12(2), 35-48. https://doi.org/10.11648/j.ab.20241202.11
ACS Style
Steve, N. S. F.; Adji, M. B.; Alphonse, S. T.; Augustin, M.; Leopold, T. N. Isolation and Probiotic Characterization of Lactic Acid Bacteria from Kindirmou and Pendidam in Adamawa Region (Cameroon). Adv. Biochem. 2024, 12(2), 35-48. doi: 10.11648/j.ab.20241202.11
AMA Style
Steve NSF, Adji MB, Alphonse ST, Augustin M, Leopold TN. Isolation and Probiotic Characterization of Lactic Acid Bacteria from Kindirmou and Pendidam in Adamawa Region (Cameroon). Adv Biochem. 2024;12(2):35-48. doi: 10.11648/j.ab.20241202.11
@article{10.11648/j.ab.20241202.11,
author = {Nodem Sohanang Francky Steve and Mohammadou Bouba Adji and Sokamte Tegang Alphonse and Mbawala Augustin and Tatsadjieu Ngoune Leopold},
title = {Isolation and Probiotic Characterization of Lactic Acid Bacteria from Kindirmou and Pendidam in Adamawa Region (Cameroon)},
journal = {Advances in Biochemistry},
volume = {12},
number = {2},
pages = {35-48},
doi = {10.11648/j.ab.20241202.11},
url = {https://doi.org/10.11648/j.ab.20241202.11},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.ab.20241202.11},
abstract = {The microorganisms intended for use as probiotics in food formulation should exert health benefit effects and be regarded as safe for animals and humans uses. The aim of this study was to evaluate the probiotic potential of lactic acid bacteria (LAB) isolated from pendidam and kindirmou, two traditional fermented milks (TFM) produced in the Adamawa region (Cameroon). Twenty-five samples (pendidam: 13 and kindirmou: 12) were randomly collected in five markets of Ngaoundere (n = 17 samples) and Meiganga (n = 8 samples). These samples were screened for their antimicrobial activity, and nine TFMs were retained. Lactic acid bacteria were isolated from these samples and their antimicrobial activity was already evaluated. Based on the inhibition zone, twenty-two LABs were retained and examined in vitro for potential probiotic properties based on their low pH tolerance, resistance to bile salts, tolerance to simulated gastrointestinal juices, hydrophobicity, autoaggregation, gelatinase and hemolytic activities. The outcome of these parameters studied was used as input data for a principal component analysis (PCA) to select the most promising isolate, and the six potential probiotic isolates were characterized through a biochemical profile. The characterized isolates have been identified as Lactiplantibacillus plantarum, Lacticaseibacillus casei, and Lactococcus lactis. Traditional fermented milks contain LAB with important properties that can be utilized in the formulation of functional foods.},
year = {2024}
}
TY - JOUR T1 - Isolation and Probiotic Characterization of Lactic Acid Bacteria from Kindirmou and Pendidam in Adamawa Region (Cameroon) AU - Nodem Sohanang Francky Steve AU - Mohammadou Bouba Adji AU - Sokamte Tegang Alphonse AU - Mbawala Augustin AU - Tatsadjieu Ngoune Leopold Y1 - 2024/04/12 PY - 2024 N1 - https://doi.org/10.11648/j.ab.20241202.11 DO - 10.11648/j.ab.20241202.11 T2 - Advances in Biochemistry JF - Advances in Biochemistry JO - Advances in Biochemistry SP - 35 EP - 48 PB - Science Publishing Group SN - 2329-0862 UR - https://doi.org/10.11648/j.ab.20241202.11 AB - The microorganisms intended for use as probiotics in food formulation should exert health benefit effects and be regarded as safe for animals and humans uses. The aim of this study was to evaluate the probiotic potential of lactic acid bacteria (LAB) isolated from pendidam and kindirmou, two traditional fermented milks (TFM) produced in the Adamawa region (Cameroon). Twenty-five samples (pendidam: 13 and kindirmou: 12) were randomly collected in five markets of Ngaoundere (n = 17 samples) and Meiganga (n = 8 samples). These samples were screened for their antimicrobial activity, and nine TFMs were retained. Lactic acid bacteria were isolated from these samples and their antimicrobial activity was already evaluated. Based on the inhibition zone, twenty-two LABs were retained and examined in vitro for potential probiotic properties based on their low pH tolerance, resistance to bile salts, tolerance to simulated gastrointestinal juices, hydrophobicity, autoaggregation, gelatinase and hemolytic activities. The outcome of these parameters studied was used as input data for a principal component analysis (PCA) to select the most promising isolate, and the six potential probiotic isolates were characterized through a biochemical profile. The characterized isolates have been identified as Lactiplantibacillus plantarum, Lacticaseibacillus casei, and Lactococcus lactis. Traditional fermented milks contain LAB with important properties that can be utilized in the formulation of functional foods. VL - 12 IS - 2 ER -
Department of Food Engineering and Quality Control, University Institute of Technology, Ngaoundere, Cameroon
Department of Food Engineering and Quality Control, University Institute of Technology, Ngaoundere, Cameroon
Department of Food Engineering and Quality Control, University Institute of Technology, Ngaoundere, Cameroon
Department of Food Sciences and Nutrition, National School of Agro-Industrial Sciences, Ngaoundere, Cameroon
Department of Food Engineering and Quality Control, University Institute of Technology, Ngaoundere, Cameroon
Information